Metamorfoza ruchu u płazów
Jednym z układów o największych zmianach podczas metamorfozy Anura jest układ lokomotoryczny. Zmiany te doprowadzają do przejścia z wodnych sposobów poruszania się na lądowe. Proces ten jest konsekwencją dynamicznej przebudowy układu nerwowego. Z rdzenia kręgowego wychodzą nowe neurony tworząc neuromotoryczną sieć obsługującą nowo wytworzone kończyny. W tym samym czasie układ unerwiający ogon ulega uwstecznieniu, a ogon zostaje zresorbowany. Ruchy ogona u larwy są napędzane poprzez dwie przyosiowe grupy mięśni, inicjujące ruchy o częstotliwości 3-5 Hz. Podczas metamorfozy utrwalają się synchroniczne, naprzemianległe ruchy kończyn o częstotliwości ok. 1-2 Hz. W czasie metamorfozy następuje zmiana charakteru impulsów wytwarzanych przez rdzeń kręgowy (CPGs) i dochodzi do sytuacji, gdy wspólnie koegzystują 2 rodzaje impulsów ruchowych - dla ogona i kończyn. Rozwój ruchowy kończyn przechodzi 3 etapy.
- 1. Jednoczesne ruchy kończynami zostają zmienione na naprzemianległe.
- 2. Częstotliwość kopnięć kończyn obniża się.
- 3. Całkowite uniezależnienie się sieci unerwiającej kończyny od układu ruchowego ogona.
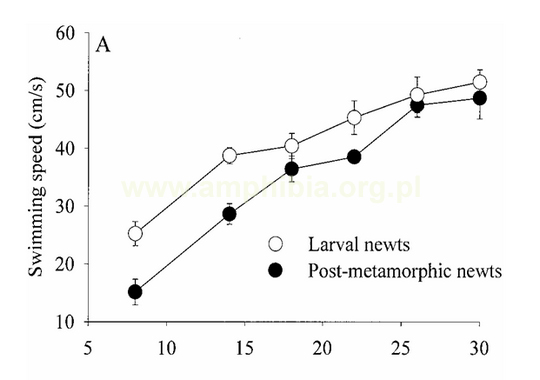
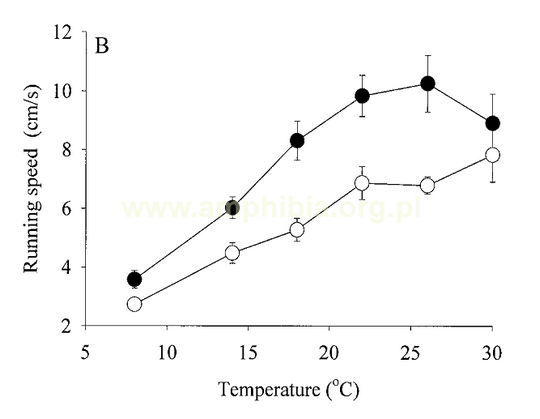
Podczas metamorfozy u Urodela zmiany układu lokomotorycznego polegają na częściowej redukcji płetwy ogonowej. Osobniki po metamorfozie potrafią poruszać się znacznie efektywniej po lądzie kosztem sprawności w wodzie. Wykres przedstawia zależność poruszania się larw (białe koła) i osobników po metamorfozie (czarne koła) w zależności od temperatury. Na wykresie A scharakteryzowano pływanie, na B - bieganie.
Główną rolę w niszczeniu komórek, bądź w ich gwałtownym namnażaniu podczas metamorfozy, pełni ubikwityna i jej cząsteczka sygnałowa - NO. Tlenek azotu jest syntetyzowany poprzez syntazę NO (NOS). Obecność tlenku azotu w wielu wypadkach odpowiada za rozwój i plastyczność neuronów (neurogeneza), szczególnie synaps (synaptogeneza) i odpowiada za sygnalizacje apoptozy w komórkach. Tlenek azotu może wywierać działanie na behawior płazów. W okresie przed metamorfozą NOS znajdują się w pewnych rejonach mózgu. Z chwilą rozpoczęcia metamorfozy syntaza NO zaczyna być produkowana w komórkach znajdujących się około powstających kończyn.
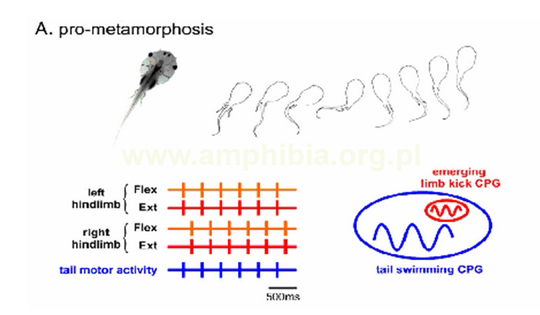
A. We wczesnych stadiach metamorfozy (pro-metamorphosis) tylne kończyny(hindlimbs) nie są jeszcze wystarczająco rozwinięte, by móc brać udział w poruszaniu się organizmu. Za pływanie odpowiedzialne są ciągle ruchy ogona, podczas gdy tylne kończyny są trzymane przy ciele. Impulsy rdzenia kręgowego, dochodzące do ogona i kończyn są ściśle skoordynowane. W czasie tego okresu, przyszły charakter CPG odpowiedzialny za ruchy kończyn osadza się w sieci nerwowej ogona (emerging limb kick CPG).
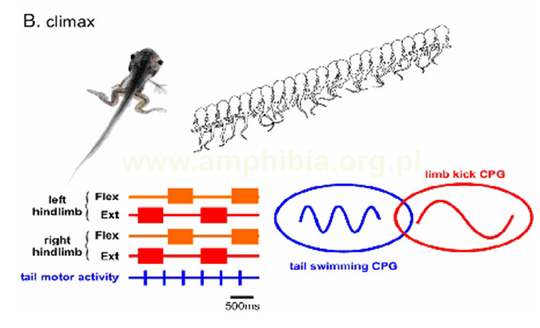
B. W punkcie kulminacyjnym metamorfozy(climax), płazy używają obu mechanizmów lokomotorycznych - ogona i kończyn, wspólnie lub niezależnie od siebie. W porównaniu z szybkimi ruchami lewa-prawa ogona (tail swiming CPG; tail motor activity), rytm ruchów kończyn tylnich jest wolniejszy i zsynchronizowany (limb kick CPG; left, right hindlimb). Ruchy mięśni kończyn tylnich - zginaczy (flexterior) i rozciągaczy (exterior), każdej kończyny są naprzemian aktywne. W tym stadium rozwoju rdzeń kręgowy generuje oddzielne impulsy nerwowe, odpowiednie dla obu mechanizmów lokomotorycznych, sugerując istnienie dwóch niezależnych motorycznych sieci nerwowych.
Publikacja: 04.11.2008 Autor: Mateusz Motyl Źródła:
"Neuromodulation and developmental plasticity in the locomotor system of anuran amphibians during metamorphosis." Sillar K., Combes D., Ramanathan S., Molinari M., Simmers J. Brain; research revives. 2007 vol. 57, str. 94-102;
"Consequences of Metamorphosis for the Locomotor Performance and Thermal Physiology of the Newt Triturus cristatus" Wilson R.; Physiological and Biochemical Zoology. 2005 vol. 75, str. 967-975;